Сформулированная выше гипотеза утверждает, что у врожденных правшей выделенная статическая информация локализована в правом полушарии, а выделенная динамическая информация — в левом. Таким образом, одно полушарие — это сознательная составляющая модели «А», а другое полушарие — подсознательная. У правшей– статиков сознательная составляющая локализована в правом полушарии, а подсознательная — в левом полушарии, у правшей– динамиков сознательная составляющая локализована в левом полушарии, а подсознательная — в правом.
Однако можно выдвинуть дополнительное предположение. Как писал один из создателей модульной теории Маунткасл:
«Каждый нейронный модуль мозга представляет собой локальную нервную сеть, которая обрабатывает информацию, передает её со своего входа на выход и при этом подвергает её трансформации, определяемой общими свойствами структуры и её внешними связями. Модули объединяются в структуры — например, в ядре или в области коры — общей или доминирующей связью, потребностью в наложении функции на определённое топографическое представительство или каким-нибудь иным фактором. Группа модулей, составляющая структуру, сама может быть разбита на подгруппы фазными связями с обособленными таким же образом подгруппами в других крупных структурах. Так, например, повсеместно полагается тот факт, что каждый корковый модуль связан с несколькими другими корковыми модулями в этом же полушарии и с зеркальным фокусом в противоположном полушарии . Этот зеркальный фокус, в свою очередь, посредством межполушарных связей связан с исходным модулем».
Логично предположить, что зеркально в коре больших полушарий расположены элементы, имеющие похожую информационную направленность. Другими словами, информация по таким информационным элементам, как ![]() и,
и, ![]() и,
и, ![]() и,
и, ![]() и, расположена симметрично в левом и правом полушарии мозга относительно мозолистого тела. Системы фильтрации информации на начальном этапе её обработки разводят информацию по полушариям мозга, однако позднее, во время её использования происходит взаимный обмен с подкачкой информации из сходных в информационной направленности сознательной и подсознательной компонент через спаечные образования мозга, т. е. через мозолистое тело, переднюю, заднюю спайки и спайки между системами свода, спаечные системы на диэнцефально–мезенцефальном уровне мозга.
и, расположена симметрично в левом и правом полушарии мозга относительно мозолистого тела. Системы фильтрации информации на начальном этапе её обработки разводят информацию по полушариям мозга, однако позднее, во время её использования происходит взаимный обмен с подкачкой информации из сходных в информационной направленности сознательной и подсознательной компонент через спаечные образования мозга, т. е. через мозолистое тело, переднюю, заднюю спайки и спайки между системами свода, спаечные системы на диэнцефально–мезенцефальном уровне мозга.
Предварительная автономность в работе полушарий мозга (сознательная и подсознательная составляющие) проявляется в том, что после перерезания мозолистого тела мозг в общем и целом способен нормально функционировать. Связь между полушариями восстанавливается через другие коммисуральные связи и подкорковые структуры. Операция не влечет за собой изменений личности, интеллекта, эмоциональной сферы, мышления, речи, заметных вне специальных условий эксперимента. «Человек, перенесший такую операцию, кажется совершенно нормальным» (Zindsay, Norman, 1972). Однако изучение больных после операции, сводящейся к перерезке анатомической связи между полушариями, принесло также данные, согласно которым для нормального функционирования и целостной психической деятельности, поддержания эмоциональных и личностных особенностей совершенно необходима передача информации из одного полушария в другое. При перерезке спаечного аппарата эта передача нарушается, «две половины мозга, не имевшие никаких секретов друг от друга» (Gazzaniga, 1970) превращаются в две функционально разобщенные системы (Мосидзе, Абкардия, 1973). В данном случае сознательный компонент лишен доступа к своему смежнику (подсознательному компоненту) (![]() и,
и, ![]() и,
и, ![]() и,
и, ![]() и
и ![]() ), а также доступа к другим информационным сегментам из полушария с противоположной установкой.
), а также доступа к другим информационным сегментам из полушария с противоположной установкой.
Подчеркнем ещё раз роль спаечного аппарата.
Чем выше ступень эволюционного развития существа, тем выше каллозно–бульбарный индекс (Anthony, 1951), отражающий соотношение площадей мозолистого тела и поперечного сечения продолговатого мозга и говорящий о взаимодействии двух полушарий. У человека этот индекс выше, чем у высших обезьян. Сопоставление площадей сечения основных спаек мозга с поверхностями кортикальных отделов анализаторов позволило установить, что развитие мозолистого тела коррелирует с развитием неокортекса (Блинков, 1964).
Def Шизофрения — психическое заболевание, которое протекает с быстрыми или медленно развивающимися изменениями личности особого типа. Такими, как снижение энергетического потенциала, эмоциональное оскудение, дискордантность (отсутствие единства). Развитие такого личностного дефекта связано с различными продуктивными синдромами и симптомами (неврозоподобными, психо-патоподобными, кататоническими, аффективными, бредовыми и т. д.) (справочник по психиатрии под редакцией Снежневского).
К. Г. Юнг о шизофрении: «В случае шизофрении вы встречаете лишь фрагменты, здесь нигде нет целого. Поэтому, если у вас есть друг или родственник, которого вы хорошо знаете и который стал сумасшедшим, вас постигнет жуткое потрясение при встрече с полностью расщепленной фрагментарной личностью» (Тэвистокские лекции).
При шизофрении значительно изменяется мозолистое тело, которое, как известно, является основным звеном анатомической основы взаимодействия между полушариями. На основе данных клинико–экспериментального исследования больных шизофренией можно говорить о признаках частичного разобщения мозга. В общей картине разобщения мозга есть проявления, расцененные, как «раздвоение личности». Например, Gazzaniga (1972) приводит описание больного с «расщепленным мозгом», который обнаруживал, что спускает брюки одной рукой и подтягивает их другой; «однажды левой рукой схватил свою жену и начал её сильно трясти, а правой рукой пытался помочь жене усмирить агрессивную левую руку». Такие факты наводят на мысль о том, что шизофрения в некоторых случаях может оказаться результатом конфликта «между двумя психическими сферами внутри одного черепа», что является ни чем иным, как конфликтом левого и правого полушарий. В этом случае, обрабатывая информацию, каждое из полушарий мозга не дополняет, а блокирует работу другого, противопоставляет свою работу работе другого полушария. Раздвоение личности при шизофрении вызывается, скорее, не нарушением системы фильтрации при обработке получаемой мозгом информации, а нарушением взаимодействия сознательной и подсознательной составляющей психики уже после получения информации.
Def Олигофрения — врожденное или рано приобретенное умственное недоразвитие, слабоумие, выраженное в недоразвитии всей психики, особенно интеллекта. Олигофрения делится на три вида недоразвития: идиотия, имбецильность, дебильность. Заболевание связано с нарушением онтогенеза, это — однородная группа заболеваний, включающая два обязательных признака: психическое недоразвитие с преобладанием интеллектуальной недостаточности и отсутствие прогридиентности (справочник по психиатрии под редакцией Снежневского).
Можно рассматривать олигофрению (умственное недоразвитие) как недоразвитие мозга, которое выражается в недоразвернутости функциональной асимметрии его полушарий. И действительно, экспериментально установлено, что чем более отстают дети в умственном развитии, тем меньше у них величина коэффициента асимметрии полушарий мозга. Существует прямая зависимость между степенью выраженности асимметрии работы полушарий мозга и умственным развитием ребёнка. Для того, чтобы накапливать опыт, необходима специализация мозга; в противном случае человек напоминает буриданова осла между двумя стогами сена. Следствием отсутствия избирательности мозга в приеме и переработке информации является слабое умственное развитие.
Гипотеза № 2.
Анализ физиологических данных по обработке зрительной информации в экстрастриарной коре позволяет выдвинуть гипотезу о том, что обработка рациональных аспектов (![]() ,,,
,,,![]() ) происходит в нижневисочной коре, а обработка иррациональных аспектов (
) происходит в нижневисочной коре, а обработка иррациональных аспектов (![]() ,
,![]() ,,
,, ![]() ) происходит в заднетеменной коре мозга.
) происходит в заднетеменной коре мозга.
Обоснование гипотезы:
Def Инвариантность — свойство объекта оставаться неизменным по отношению к некоторым его преобразованиям.
На основе ряда клинических и физиологических фактов была выдвинута гипотеза о раздельной локализации в коре головного мозга двух механизмов описания зрительных изображений. Механизм классификации локализован у приматов в нижневисочной коре ( рациональная компонента), а механизм конкретного описания изображений — в заднетеменной коре ( иррациональная компонента) (Глезер, 1978, 1985). Реакция на изображения определяется величиной вклада двух одновременно действующих механизмов — механизмом инвариантного изображения (нижневисочная область) и механизмом оценки варианта (заднетеменная область).
Некоторые краткие данные для нижневисочной коры (локализация,,, ![]() ):
):
Обезьяны без нижневисочной коры быстро обучаются различать стимулы одной формы, но разной ориентации и размера (Holms, Gross, 1984) (т. е. у них сохраняется восприятие иррациональных компонент при утраченной способности работать по рациональным компонентам). У животных, оперированных в нижневисочной области, понижаются показатели в обучении зрительным дифференцировкам формы и сохранению дооперационных дифференцировок ( рациональные компоненты).
Некоторые краткие данные для заднетеменной коры (локализация,,, ![]() ):
):
Разрушение заднетеменной коры не влияет на сохранность дифференцировок, выработанных до операции и на образование новых (т. е. обработка рациональных компонент идёт по-прежнему успешно). В то же время, обезьяны без заднетеменной коры хуже опознают одни и те же преобразованные стимулы, чем интактные животные, т. к. на уровне заднетеменной коры локализован механизм, который оценивает пространственные признаки (в частности, ориентацию и размер). Изображения одинаковой формы, но отличающиеся размером и ориентацией, на уровне заднетеменной коры имеют существенно различающиеся описания.
Оцениваемые признаки в нижневисочной коре: дифференцировка по форме независимо от цвета, размера, яркости, ориентировки, выделения так называемых выпуклых признаков. По форме (схеме) объект классифицируется.
Оцениваемые признаки в заднетеменной коре: обобщение таких свойств реального мира, как цвет, ориентация, размер, яркость. По ориентации и цветовому окрашиванию в данный момент времени у объекта выделяются характерные черты.
Таким образом, в зрительной системе осуществляется, с одной стороны, инвариантное описание зрительных объектов — формируется зрительный образ (некоторая схема, т. е. рациональные аспекты), с другой — дается описание отдельных свойств этого объекта, обеспечивающее конкретизацию зрительного образца (ситуативные нюансы, т. е. иррациональные аспекты).
Вывод на основе данных по очаговым поражениям мозга:
На основе гипотез № 1 и № 2 можно привести схему мозга с локализацией механизмов обработки аспектов по левому и правому полушариям и ассоциативным зонам нижневисочной и заднетеменной коры. А именно, рациональные, статические аспекты (![]() и
и ![]() ) обрабатываются в правой нижневисочной коре, рациональные динамические аспекты (
) обрабатываются в правой нижневисочной коре, рациональные динамические аспекты (![]() и
и ![]() ) обрабатываются в левой нижневисочной коре, иррациональные статические аспекты (
) обрабатываются в левой нижневисочной коре, иррациональные статические аспекты (![]() и
и ![]() ) обрабатываются в правой заднетеменной коре, а иррациональные динамические аспекты (
) обрабатываются в правой заднетеменной коре, а иррациональные динамические аспекты (![]() и
и ![]() ) обрабатываются в левой заднетеменной коре.
) обрабатываются в левой заднетеменной коре.
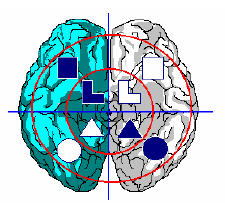
Рис. 3. Локализация аспектов в долговременной памяти.
Человек усваивает информацию разных модальностей, однако 90% информации, поступающей в мозг, воспринимается с помощью глаз (Lindgren N., 1962). Через работу зрительной системы человека можно выявить фильтрационные механизмы на конкретной стадии обработки информации. Отслеживая то, что происходит на той или иной стадии, мы, возможно, поймем, как преобразуется входной зрительный сигнал и какие признаки обрабатываются на каком этапе.
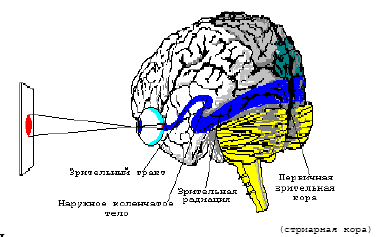
Рис. 4. Постановка эксперимента по регистрации ответов в зрительной коре.
При обработке зрительной информации выделяют 3 основных блока. А именно, на начальном этапе информация с сетчатки по зрительным нервам поступает для обработки через промежуточный и средний мозг в наружное коленчатое тело (НКТ). Затем, после НКТ, происходит обработка зрительной информации в первичной зрительной коре (стриарная кора), после чего обработанная зрительная информации поступает в экстрастриарную кору (престриатум). Проследить дальнейшее движение зрительной информации практически невозможно.
Схематически этот путь можно изобразить следующим образом:
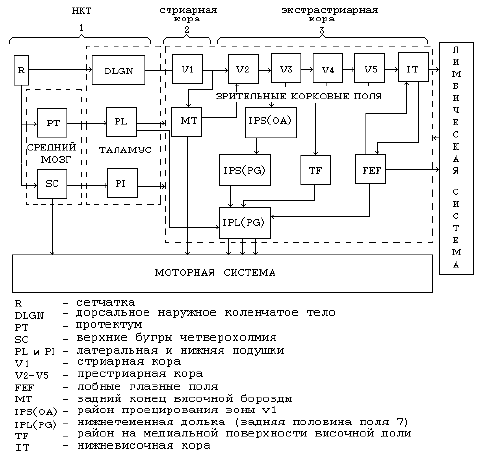
Рис. 5. Три основных этапа в обработке зрительной информации.
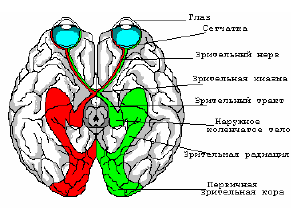
Рис. 6. Зрительные пути в мозгу человека.
Тахистоскопический принцип подачи сигналов таков, что объекты, находящиеся в левом поле зрения, раздражают правую половину сетчатки каждого глаза, нервные волокна из этих частей сетчатки обоих глаз приходят в зрительную кору правого полушария; зрительная информация из правого поля зрения поступает в левое полушарие.
Следовательно, такой индикатор, как расположение относительно объекта исследования, возможно, сработает для определения признака статика – динамика.
Гипотеза: правша– статик предпочитает находиться относительно объекта исследования справа от него, а правша– динамик — слева от него.
Рассмотрим подробнее три этапа обработки информации.
В основу гипотезы Хьюбела и Визела о функциональной организации зрительной коры положены два принципа: детекторный принцип обработки сигнала и иерархический принцип построения системы детекторов. Детекторный принцип позволил обнаружить, что нейроны зрительной коры выделяют определённые признаки изображения. Иерархический принцип говорит о том, что нейроны с рецептивными полями (простые–сложные–сверхсложные) представляют собой последовательные, все более высокие стадии обработки зрительной информации, на которых выявляются все более сложные признаки изображения.
Первый уровень обработки зрительной информации — это описание изображения множеством рецептивных полей зрительной подкорки. Нейроны сетчатки и наружного коленчатого тела измеряют интегрированную световую энергию в пределах центральной зоны суммации рецептивных полей. Существование тормозной периферии рецептивных полей приводит к тому, что нейроны этого уровня выполняют ряд операций по предварительной переработке зрительного сигнала: выделение сигнала из шума, подчеркивание контуров и высоких пространственных частот, пространственную и временную декорреляцию изображения. Ретинотопическая и структурная упорядоченность в организации наружного коленчатого тела, а также выявленные у рецепторных нейронов свойства позволяют предположить существование на этом уровне зрительной системы квазипоточечного описания зрительного мира, т. е. отображение в пространственно–временном рельефе активности нейронов наружного коленчатого тела всех свойств изображения, существующего на сетчатке. Пространственные характеристики такого рельефа активности несут в себе сведения о пространственных свойствах изображения и освещенности изображения; т. е. предполагается, что подкорковые уровни служат для эффективного кодирования, обработки и передачи в кору сведений об отдельных параметрах ретинального изображения (интенсивность, размер, элементы формы и т. д.).
Таким образом, на уровне первичной обработки информации в сетчатке и на уровне обработки информации в наружном коленчатом теле происходит подготовка информации (своего рода выделение сигнала из шума) для её дальнейшей фильтрации в стриарной и экстрастриарной коре.
Рецептивные поля стриарной коры (поле 17 или V1) — двумерные решетчатые фильтры пространственных частот. Поля измеряют пространственно–частотную и ориентационную компоненты фрагментов изображения. Нелинейные нейроны служат для сегментации зрительного поля на отдельные фрагменты. Для каждого отдельного фрагмента эта операция может быть описана как выделение фигуры из фона. Выполняется она сложными нейронами, измеряющими кусочные мощностные спектры и выделяющими участки, занятые однородной текстурой; дирекциональными нейронами, выделяющими площадки, составленные элементами, движущимися в одном направлении; бинокулярными нейронами, выделяющими участки, расположенные на разной глубине, а также цветовыми нейронами. Эти участки могут быть объединены в более крупную фигуру обратными влияниями из вышележащих отделов. Выделение участка анализируется модулем, составленным линейными нейронами. Каждый модуль состоит из рецептивных полей одной величины и разной ориентации. Для каждого выделенного изображения подбирается модуль соответствующего размера и положения из сети модулей, перекрывающей центральный участок поля зрения. Показано, что такая система модулей может быть отождествлена с оптимальной системой передачи информации по Габору, поскольку все модули могут быть соотнесены с логонами, имеющими одинаковую площадь. Модель модулей объединяет концепции модулей Маунткасля, ориентационных свойств Хьюбела–Визела и пространственные частоты каналов Кэмпбелла.
Таким образом, легко заметить, что в стриарной коре происходит выделение информационных фрагментов по признаку фигура–фон (тело–поле, объект–взаимодействие (окружение)). Акцент делается либо на выделении фигуры из фона, либо на выделении фона у фигуры.
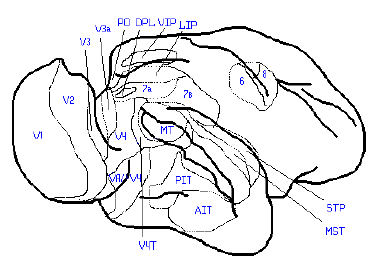
Жирными линиями отмечены границы мозга и основные борозды.
Тонкими линиями отмечены примерные границы зрительных зон.
Рис. 7. Расположение зрительных зон в коре мозга обезьяны макаки.
Экстрастриарный пояс в коре обезьян и человека — это зоны V2, V3, VP, V4, V4T, MT = V5, MST, VIP, V3A, PO, DPL.
Опишем работу нейронов в данных зонах экстрастриарного пояса:
V2: Возбуждение нейронов зоны при определённом положении стимула по глубине относительно точки фиксации; реагирование на скорость движения стимула (предпочтение медленной скорости в центре и быстрой скорости на периферии), избирательность к направлению движения, цветовая чувствительность и ориентационная избирательность положения стимула.
V3 и VP: По нейронному составу в зоне V3 доминируют нейроны с дирекциональной избирательностью, в зоне VP — цветоизбирательные нейроны.
V4, VA/V4, V4T: Отмечены нейроны с дирекциональной и ориентационной избирательностью (движение стимулов), избирательностью к определённой длине стимула и чувствительностью к текстурным поверхностям. В V4 и V4T существуют также нейроны с цветовой чувствительностью.
V5==MT: Особая чувствительность к движению (интервал скоростей от 2 до 256 гр/с). Многие нейроны проявляют избирательность к направлению движения и к скорости. Часть нейронов, кроме того, настроена на определённые диспаратности проекций стимулов на сетчатку глаз, т. е. избирательны к положению стимула по глубине относительно точки фиксации. В зоне MT описаны нейроны, избирательно реагирующие на сближение или удаление определённым образом ориентированных границ в пределах рецептивного поля.
MST и VIP: У зоны MST наиболее сильные ответы вызываются небольшими резкими ответами мелких стимулов, существует избирательная реакция на увеличение или уменьшение зрительного стимула и на определённое направление вращения стимула как во фронтальной плоскости, так и в глубину. Свойства ответов нейронов в зоне VIP пока исследованы не были.
V3A, PO и DPL: Зона V3A расположена в пограничной области между затылочной и теменной корой. У нейронов зоны V3A замечена интересная особенность: величина их ответов на одинаковую стимуляцию сетчатки в значительной степени менялась при изменении направления взора животного. Всесторонние исследования зон PO и DPL пока не проводились.
Экстрастриарные зрительные зоны, описанные выше, большей частью располагаются в отделах коры, относимых к ассоциативным.
Таким образом, мы видим, что в экстрастриарной коре анализируются движущиеся или покоящиеся текстуры, т. е. работает фильтрационный механизм признака статика – динамика.
После зон экстрастриарного пояса информация поступает в основном в две области: в нижневисочную кору и в заднетеменную кору. По существу, к этому моменту за счёт фильтрационных механизмов потоки в нижневисочную кору несут аспекты,,,, в заднетеменную кору — аспекты,,, ![]() .
.
Примечательно, что именно в этих двух основных высших отделах зрительной системы — нижневисочном и заднетеменном, содержатся нейроны, активность которых коррелирует с обучением. Предполагается, что обучение осуществляется наиболее простым, рекуррентным способом, что возможно благодаря оппонентной организации на предшествующих уровнях.
Информация по этим двум отделам приводилась выше. Сейчас мы её несколько дополним.
Зрительные зоны височной коры обезьян участвуют в обучении зрительным стимулам. При поражении данного участка коры обезьяны необратимо теряли способность обучаться различению зрительных стимулов ( рациональные аспекты), в то же время они имели сохранное поле зрения, нормальную разрешающую способность и не отличающиеся от нормы пороги ряда зрительных психофизиологических функций.
Одновременно с нижневисочным работает заднетеменной отдел зрительной системы. Его задача — конструировать экстра- и интроперсональное пространство с помощью механизмов, называемых фреймами. В процессе зрительного акта ячейки фрейма заполняются с помощью механизма избирательного внимания. Запрос направляется по обратной связи на любой уровень зрительной системы и позволяет получить развернутую информацию об участках зрительного поля или о компонентах его спектрального описания. Благодаря этому происходит съём нужной информации, в том числе и о пространственных отношениях. Получение полной информации о пространственных отношениях между объектами или между элементами объекта равноценно их полному описанию (конкретизации).
Анализ лингвистических концепций и гипотез теории искусственного интеллекта подтверждает, что парадигматическую и синтагматическую функции языка можно сопоставить с двумя исследованными в зрительной коре механизмами. Для парадигматической функции базисом на сенсорном уровне является механизм классификации, работающий с помощью разделительных признаков (словарь, иррациональная обработка), синтагматическая функция языка основывается на механизме описания отношений (грамматике, рациональная обработка). Таким образом, в языке в виде его парадигматической и синтагматической функций отражаются обе главные функции сенсорного мозга: классификация образов и конкретное описание. Упорядоченное хранение информации и быстрый поиск нужных кодов достигаются благодаря тому, что в конструкции мозга заложены две основные формы организации языка: словарь и грамматика.
Вывод на основе физиологических данных по обработке зрительной информации:
При прохождении информации от сетчатки до проекционных зон больших полушарий последовательно срабатывают фильтрационные механизмы. На этапе первичной обработки информации и прохождения через наружное коленчатое тело информация готовится для первой стадии фильтрации в стриарной коре. На втором этапе (стриарная кора) непосредственно запускается механизм фильтрации по признаку фигура–фон (тело–поле). На третьем этапе (этапе прохождения информации через экстрастриарную кору) работает фильтрационный механизм по признаку статика – динамика . Далее, на этапе попадания информации в нижневисочную и заднетеменную кору происходит обращение к эталону признака внешнее – внутреннее. После этапа фокусировки слоя сознания, по всей видимости, результирующая информация после прохождения всех распознающих фильтрационных механизмов собирается в единый образ.
Нами установлено месторасположение механизмов обработки аспектов в коре головного мозга и стадиальность включения фильтров. Наша задача теперь — проанализировать эти данные и попытаться вывести из них модель «А». Для этого, по мере анализа информации, будем выдвигать постулаты, которые наиболее естественным образом описывают наблюдаемые явления.
Говорить о том, что одно полушарие только «статично», а второе только «динамично» неверно, так как в левом «динамическом» полушарии присутствует информация и по статическим аспектам, но на актуальном базисе после нескольких фильтров практически оседает информация только по динамическим аспектам, в правом полушарии — практически только по статическим аспектам.
Предполагается, что полная информация (т. е. информация обо всех 8 аспектах) поступает на оба глаза. От левых полей зрения информация поступает в левое полушарие, от правых полей зрения — в правое полушарие. Заметим, что как в левом, так и в правом полушариях содержится информация обо всех 8 аспектах.
Постулат № 1: На уровне стриарной коры определена структура, доминантная по одному из полюсов фигура–фон. Причем полюс доминантности в одном полушарии прямо противоположен полюсу доминантности в другом полушарии.
Например, пусть в левом полушарии закреплена доминантность по полюсу фон. Тогда в правом полушарии будет закреплена доминантность по полюсу фигура . Это приведет к тому, что вся информация в левом полушарии профильтруется таким образом, что будет выделен фон (![]() ,,,
,,, ![]() ), сами фигуры (
), сами фигуры (![]() ,,,
,,, ![]() ) станут «фоновой» информацией по отношению к этому фону . В правом же полушарии, наоборот, информация профильтруется таким образом, что будут выделены фигуры (
) станут «фоновой» информацией по отношению к этому фону . В правом же полушарии, наоборот, информация профильтруется таким образом, что будут выделены фигуры (![]() ,,,
,,, ![]() ), сам же фон (
), сам же фон (![]() ,,,
,,, ![]() ) останется «фоном» по отношению к выделенным фигурам .
) останется «фоном» по отношению к выделенным фигурам .
Здесь же, по всей видимости, включается механизм доминирования в обработке информации одного полушария (признак ментальность–витальность у А. Аугустинавичюте).
Постулат № 2: На уровне экстрастриарной коры определена структура, доминантная по одному из полюсов статика – динамика . Причем полюс доминантности в левом полушарии — динамика, в правом полушарии — статика.
Например, следуя вышеприведенному выбору, информация в левом полушарии профильтруется таким образом, что будут выделены динамические аспекты (![]() ,
, ![]() ) на фоне статических (
) на фоне статических (![]() ,
, ![]() ) и (
) и (![]() ,
, ![]() ) на фоне (
) на фоне (![]() ,
, ![]() )). В правом же полушарии, наоборот, информация профильтруется таким образом, что будут выделены статические аспекты (
)). В правом же полушарии, наоборот, информация профильтруется таким образом, что будут выделены статические аспекты (![]() ,
, ![]() ) на фоне динамических (
) на фоне динамических (![]() ,
, ![]() ) и (
) и (![]() ,
, ![]() ) на фоне (
) на фоне (![]() ,
, ![]() ).
).
По всей видимости, существуют врожденные структуры, отвечающие за приоритетное направление в передаче информации (от нижневисочных к заднетеменным отделам коры больших полушарий или наоборот). Эти структуры приводят к разному характеру работы нижневисочного и заднетеменного отделов мозга (один из них акцептен, другой — продуктивен ), что на уровне модели «Ю» выражается в существовании у человека только одной, cамой сильной функции (это та функция, которая постоянно копит информацию), а на уровне модели «А» выражается в появлении, например, самой сильной функции (1) и вспомогательной (2) (заметим, что эти функции самые сильные у типа в обработке информации, работа по этим функциям хорошо осознается), а также аналогичным раскладкам по остальным блокам (помимо ЭГО).
Т. е. через доминирование одного из отделов мозга (нижневисочного или заднетеменного) «включается» работа признака акцептность – продуктивность у А.Аугустинавичюте.
Постулат № 3: На уровне нижневисочной коры определена структура, отвечающая за уровень актуального базиса по оси внешнее–внутреннее. На уровне заднетеменной коры определена аналогичная структура. Причем уровень актуального базиса на ней может отличаться от уровня в нижневисочной коре.
Здесь стоит сделать некоторое отступление. Что такое признак внешнее – внутреннее? Это — месторасположение эталона, по которому мы оцениваем приходящую информацию. В случае внешнего эталона рассматриваемый объект объективно оценивается по внешним критериям (цена за товар определённого качества (![]() ), наблюдаемая работа (
), наблюдаемая работа (![]() ), степень соответствия или несоответствия предлагаемому извне закону (например, судебному —
), степень соответствия или несоответствия предлагаемому извне закону (например, судебному — ![]() ), внешне оцениваемая эстетика интерьера (
), внешне оцениваемая эстетика интерьера (![]() )). В случае внутренней оценки, эталон находится глубоко внутри нас. И поскольку этот эталон внешне не наблюдается, то в этом случае мы можем говорить не об объективной, а о субъективной оценке. При этом степень точности соответствующих оценок объектов у последнего отнюдь не ниже, чем у первого, просто она имеет другую природу (внутреннюю). Внутренний эталон проявляется в проницательности при оценивании потенциала человека (
)). В случае внутренней оценки, эталон находится глубоко внутри нас. И поскольку этот эталон внешне не наблюдается, то в этом случае мы можем говорить не об объективной, а о субъективной оценке. При этом степень точности соответствующих оценок объектов у последнего отнюдь не ниже, чем у первого, просто она имеет другую природу (внутреннюю). Внутренний эталон проявляется в проницательности при оценивании потенциала человека (![]() ), тонком внутреннем ощущении своевременности или несвоевременности тех или иных событий (
), тонком внутреннем ощущении своевременности или несвоевременности тех или иных событий (![]() ), успехе работы психотерапевта над изменением отношения клиента к своей проблеме (
), успехе работы психотерапевта над изменением отношения клиента к своей проблеме (![]() ), артистических способностях человека, умении точно выражать внутреннее эмоциональное состояние (
), артистических способностях человека, умении точно выражать внутреннее эмоциональное состояние (![]() ).
).
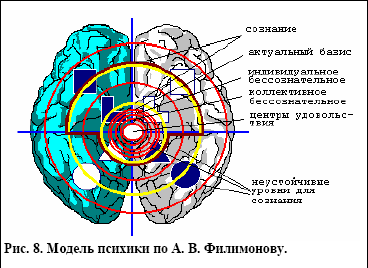
Рис. 8. Модель психики по А. В. Филимонову.
К этому моменту у нас существует четыре потока информации (в одном из них в сплавленном виде доминирует ![]() и, во втором —
и, во втором — ![]() и, в третьем —
и, в третьем — ![]() и, в четвертом —
и, в четвертом — ![]() и
и ![]() ).
).
Пока, чисто интуитивно, создается впечатление, что на этом этапе никакой третьей фильтрации не происходит и вся информация по вышеперечисленным парам остается в едином сжатом виде, однако при работе с данной информацией требуется её соотнесение с существующими эталонами (внешними или внутренними). Постоянное колебание эталонов по этой оси в той же степени, как и полное доминирование одного эталона и погашение второго, ведут к неустойчивому уровню сознания (в первом случае — это противоположные требования к информации, во втором — исключение одного из эталонов, что чревато дезориентацией в целой психической сфере). Это приводит к генетически обусловленной или формируемой на ранних этапах развития фиксации местоположения сознания в виде актуального базиса, при котором информация по одному эталону практически не искажается (в этом проявляется «сила» обрабатывающей соответствующий аспект функции) и в тоже самое время эта же информация при соотнесении со вторым эталоном значительно искажается (в этом проявляется «слабость» функции, обрабатывающей второй аспект из пары).
Таким образом, локализация актуальных базисов в нижневисочной и заднетеменной областях связана с признаком «сильная–слабая» для функций в модели А. Аугустинавичюте.
Cоционические типы можно представить схематически, исходя из вышеперечисленных четырёх неизменных на протяжении жизни отдельного человека параметров: доминирование одного из полушарий, направленность в передаче информации (направленность нейронных потоков) между двумя основными ассоциативными зонами коры больших полушарий, фиксация актуального базиса в нижневисочной коре, фиксация актуального базиса в заднетеменной коре.
(На схеме условно местоположение внешних эталонов соответствует внешнему кругу, внутренних эталонов — внутреннему кругу).

Рис. 9. Схематическое обозначение соционических типов, исходя из доминантности полушария, направленности потоков информации и фиксации актуальных базисов.
Приведенные данные не только дают физиологическую основу моделям «Ю» и «А», но и хорошо объясняют колебание в дихотомиях интуиция – ощущение, чувство – мышление, экстраверсия – интроверсия (разделение дихотомии экстраверсия – интроверсия происходит на этапе фильтрации фигура–фон, т. к. экстравертная установка подразумевает выделение фигуры, а интровертная — выделение фона) у Юнга, вскрывают основу аспектного и функционального деления у Аугустинавичюте и выводят существование только 16-ти устойчивых состояний психики в соционике.
У Аушры Аугустинавичюте рассматривалось понятие ведущего глаза. Ведущий глаз связан с сознательной установкой. Соответственно у циклотимов, по А. Аугустинавичюте, более «осознан» левый глаз, если они статики, и правый глаз, если они динамики. У шизотимов — наоборот.
Действительно, вопрос о существовании ведущего глаза заслуживает особого рассмотрения. Ведущий глаз, если он выделен, первым устанавливается на предмет, тогда как другой глаз заканчивает общую установку, направляя зрительную ось на точку фиксации ведущего глаза. В ведущем глазу раньше включается механизм аккомодации. Изображение ведущего глаза преобладает над изображением подчинённого; в ведущем глазу нейтрализуются нечёткие детали изображения подчинённого глаза. При фиксации предмета ведущий глаз управляет установкой подчинённого. У неведущего глаза мышцы развиты слабее и негармоничнее. Из 601 человека, обследованного А. Латинским, асимметрия глаз обнаружена у 92,6%, у 7,4% обнаружена симметрия — равноценность обоих глаз. Среди лиц с асимметрией правый глаз преобладает у 62,6%, левый — у 30% обследованных. Другие данные: A. Brusa, G. Francia, L. Gregiretti (1957) — правоглазые (24%), левоглазые (27%), лица без ведущего глаза (49%); В. Мосидзе (1977) — правоглазые (28,6%), левоглазые (4,8%). Видно, что статистические данные разноречивы: эти же данные не подтвердили гипотезу о том, что ведомость глаза связана с леворукостью–праворукостью.
Набранная мной статистика по ведущему глазу не только не подтвердила корреляцию ведомости глаза с предположением А. Аугустинавичюте, но и не выявила связи ни с одним из признаков Рейнина. Наличие ведущего глаза, на мой взгляд, вытекает просто из того факта, что у человека два глаза, а не один, и как следствие такой ситуации, один иногда доминирует. Оба глаза несут одинаковую функциональную нагрузку. За счёт работы парного органа достигается бинокулярная локализация объекта в трехмерном пространстве, а как следствие этого — более полный зрительный образ и лучшая координация при движении.
Выдвинутые гипотезы вносят ясность в аналитическую психологию Юнга и в соционику Аугустинавичюте, однако успехом можно считать только их экспериментальное подтверждение. Для этого создан необходимый стимульный материал (компьютерная программа, предлагающая на разных этапах различный стимульный материал в дихотомиях фигура–фон, статика–динамика и в смешанном варианте, и вызывающая фиксацию на полюсах дихотомий) и продуманы методы диагностики активированных зон мозга на электроэнцефалографе во время реагирования на стимульный материал.
Существенное достоинство этих тестов состоит в оригинальности предлагаемого стимульного материала, а также в самой специфике его подачи, при которой испытуемого не заставляют выполнять какую-то задачу, а снимают естественную реакцию мозга на тот или иной стимульный материал.
Данная работа предоставляет физиологическую основу для аналитической психологии K. Г. Юнга, его теории, определяющей психологические типы. Это же в равной степени относится и к соционике.
Модели «А», и тем более «Ю», в настоящей работе не принимаются за постулат, а выводятся на основе строения головного мозга человека. На основании этой информации выводится число соционических типов и законы обработки аспектов cоответствующими функциональными способами.
Более того, эта работа выходит за рамки просто соционических знаний. Она указывает место соционики в мире изучения психической активности человека.
Работа имеет большое практическое значение. Она не только дает необходимую теоретическую основу для первого этапа обработки информации в когнитивной психологии (сенсорно–перцептивный блок в микроструктурном анализе), но и предполагает точные физиологические методы при диагностике психологического типа.
Вполне возможно, что хорошо продуманные и организованные эксперименты вызовут лавинообразный поток исследований и повлекут за собой новые открытия в изучении человеческой психики.